El estudio de la herencia genética ha fascinado a la humanidad desde tiempos remotos. La manera en que los rasgos físicos, fisiológicos e incluso conductuales se transmiten de padres a hijos ha sido objeto de curiosidad, investigación y controversia a lo largo de la historia. Hoy en día, gracias al desarrollo de la genética como disciplina científica y al uso de técnicas de laboratorio altamente sofisticadas, comprendemos mejor los mecanismos subyacentes a esta transmisión. Sin embargo, antes de adentrarnos en la explicación detallada de los genes, los alelos y su expresión, conviene hacer un repaso histórico y conceptual que nos permita entender cómo hemos llegado a comprender la distinción entre rasgos (o características) dominantes y recesivas.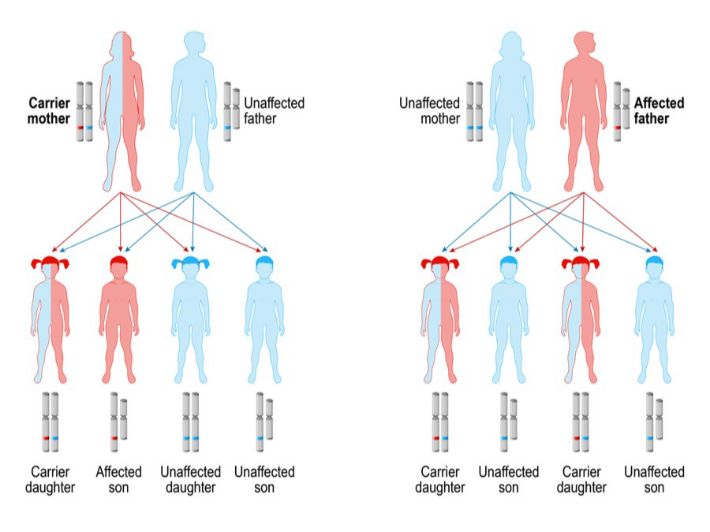
En la presente publicación se explorarán de manera extensa y detallada las distintas facetas de la herencia genética, enfocándonos en los conceptos de dominancia y recesividad, pero también ampliando la discusión hacia contextos más complejos y realidades biológicas que no siempre siguen el patrón mendeliano clásico. Con un enfoque científico y a la vez divulgativo, se buscará ofrecer información de alto valor, tanto para la comunidad académica como para lectores curiosos que deseen ampliar su conocimiento sobre este apasionante ámbito de la biología.
Se ha estructurado este artículo en varios apartados, con el fin de abarcar las diferentes perspectivas y profundizar lo suficiente en cada una de ellas. Al término del mismo, se incluyen referencias bibliográficas y una tabla ilustrativa con ejemplos concretos de rasgos dominantes y recesivos, lo que permitirá asentar los conceptos expuestos. El propósito es ofrecer una pieza de contenido que cumpla con rigor científico, pero que a la vez sea amena y de utilidad práctica para el lector.
1. Breve contexto histórico de la genética y la herencia
La idea de que los rasgos se transmiten de una generación a otra se remonta a la antigüedad. Sin embargo, durante siglos, el entendimiento de este fenómeno se basó en especulaciones y explicaciones poco fundamentadas. Filósofos y científicos de la Grecia clásica, como Aristóteles, propusieron teorías que intentaban explicar la herencia a partir de la influencia de “fluidos” o esencias que se mezclaban en la descendencia. No fue hasta el siglo XIX, con el trabajo pionero de Gregor Mendel, que se establecieron las bases cuantitativas de la herencia, marcando un antes y un después en la comprensión de los mecanismos hereditarios.
Gregor Mendel, un monje agustino, trabajó meticulosamente en el estudio de la herencia en plantas de guisante (Pisum sativum). Gracias a sus experimentos, logró inferir la existencia de “factores hereditarios” (que hoy llamamos genes) y describir cómo estos factores se transmitían en patrones de dominancia y recesividad. Para explicar sus hallazgos, postuló las Leyes de Mendel: la Ley de la Segregación, la Ley de la Distribución Independiente y la Ley de la Uniformidad de los Híbridos de la Primera Generación. Durante mucho tiempo, su obra pasó desapercibida, pero a inicios del siglo XX fue redescubierta por varios investigadores y se convirtió en la piedra angular de la genética.
Con el descubrimiento posterior de la estructura del ADN por parte de James Watson, Francis Crick, Maurice Wilkins y Rosalind Franklin, y con el desarrollo de la biología molecular a lo largo del siglo XX, quedó claro que los genes estaban constituidos por secuencias de ADN y que el modo en que se expresan determina en gran medida nuestras características biológicas. El concepto de rasgos dominantes y recesivos se ha mantenido esencial, pero también han surgido matices y excepciones que han enriquecido nuestra comprensión de la genética.
2. Conceptos clave en genética: genes, alelos y locus
2.1. Definición de gen
Un gen puede definirse como una unidad de información en el ADN que codifica un producto funcional, ya sea una proteína o un ARN con función biológica. En el genoma de un organismo, los genes están organizados de manera lineal en los cromosomas. Su función esencial es aportar la información necesaria para la síntesis de proteínas que cumplen roles estructurales, enzimáticos, reguladores y de defensa, entre otros.
2.2. Concepto de alelo
El término “alelo” se refiere a las variantes o formas alternativas de un mismo gen. Para un gen dado, pueden existir múltiples alelos en la población, aunque cualquier individuo diploide (como los seres humanos) posee tan solo dos copias de cada gen, una procedente del progenitor materno y otra del progenitor paterno. La presencia de distintas variantes alélicas es la base de la diversidad genética y explica por qué dos individuos de la misma especie pueden presentar diferencias en ciertos rasgos, como el color de ojos o el tipo de cabello.
2.3. Locus y organización cromosómica
El locus (plural: loci) es la posición específica que ocupa un gen o un marcador genético en un cromosoma. Conocer la ubicación exacta de cada gen es importante para entender cómo se heredan determinadas características y cómo pueden presentarse mutaciones que causen enfermedades. El avance de la genómica y las técnicas de mapeo han permitido cartografiar los genomas de múltiples organismos, facilitando la identificación y el estudio de los genes asociados con rasgos concretos o con patologías genéticas.
3. Dominancia y recesividad: fundamentos mendelianos
3.1. Definiciones iniciales
En términos mendelianos, un rasgo se considera dominante cuando se expresa en la descendencia incluso si solamente está presente un alelo que lo determina. Por el contrario, un rasgo recesivo únicamente se manifiesta cuando ambos alelos que posee el individuo para ese gen son recesivos. De esta manera, si un individuo diploide hereda un alelo dominante y uno recesivo, el fenotipo observado será el dictado por el alelo dominante.
Para ilustrar esto con un ejemplo clásico, en los guisantes que estudió Mendel, la forma lisa de la semilla es un rasgo dominante, mientras que la forma rugosa es recesiva. Si se cruza una planta homocigota dominante (con dos alelos dominantes) con una planta homocigota recesiva (con dos alelos recesivos), toda la primera generación filial (F1) presentará semillas lisas. No obstante, en la segunda generación filial (F2), reaparecerán plantas con semillas rugosas en una proporción de 3:1, resultado típico de la separación alélica y la combinación de los mismos en la fecundación.
3.2. Homocigosis y heterocigosis
Un individuo es homocigoto para un rasgo cuando posee dos copias idénticas de un determinado gen (ya sea AA para la forma dominante o aa para la forma recesiva). Por el contrario, es heterocigoto si posee dos alelos diferentes (Aa) para ese gen. En el caso de la heterocigosis, la expresión fenotípica vendrá determinada por el alelo dominante, asumiendo que el patrón de herencia es estrictamente mendeliano y no hay otras interacciones alélicas como la codominancia o la dominancia incompleta.
3.3. Leyes de Mendel y su pertinencia actual
Las tres leyes de Mendel constituyen el núcleo de la genética clásica, pero no abarcan todas las complejidades observables en la herencia. Aun así, siguen siendo una base fundamental para entender cómo se heredan gran parte de los rasgos y cómo se comportan los alelos en la reproducción sexual. Sus principios resultan válidos para numerosos organismos y se aplican, con matices, a humanos, animales y plantas.
- Ley de la Segregación: Los alelos de un gen se separan durante la formación de los gametos, de modo que cada gameto recibe un solo alelo.
- Ley de la Distribución Independiente: Distintos pares de alelos se distribuyen de manera independiente durante la formación de los gametos, siempre y cuando no se encuentren en el mismo cromosoma (o si se encuentran, que estén suficientemente separados para que ocurra recombinación).
- Ley de la Uniformidad: Todos los descendientes de un cruce entre progenitores homocigotos para diferentes alelos de un mismo gen presentarán un fenotipo uniforme (el del alelo dominante, en el caso de que exista dominancia completa).
4. Variaciones sobre la dominancia: más allá del modelo simple
El modelo inicial propuesto por Mendel de “un alelo dominante y uno recesivo” ha demostrado ser muy útil para describir la herencia de ciertos rasgos. Sin embargo, las investigaciones genéticas han revelado una amplia gama de fenómenos que amplían o matizan los conceptos de dominancia y recesividad. A continuación, se analizarán algunos de estos fenómenos.
4.1. Dominancia incompleta
En la dominancia incompleta, el fenotipo de los individuos heterocigotos es intermedio entre el de los homocigotos dominantes y los homocigotos recesivos. Un ejemplo clásico ocurre en ciertas variedades de flores: si se cruza una planta de flores rojas (RR) con otra de flores blancas (rr), la descendencia heterocigota (Rr) presentará un color rosado. En este caso, ninguno de los alelos domina completamente al otro y se observa una mezcla fenotípica.
4.2. Codominancia
La codominancia sucede cuando dos alelos distintos se expresan de manera simultánea en el fenotipo del heterocigoto. Un ejemplo muy conocido es el grupo sanguíneo AB en humanos, donde los alelos A y B se expresan de forma completa en la superficie de los glóbulos rojos, generando un fenotipo que exhibe las características de ambos alelos sin que uno suprima la expresión del otro.
4.3. Alelismo múltiple
En la naturaleza, es frecuente que existan múltiples variantes alélicas para un mismo gen. Esto va más allá del planteamiento mendeliano de un solo par de alelos (A y a). Un ejemplo claro es también el del sistema de grupos sanguíneos ABO, donde el gen que determina este rasgo cuenta con tres alelos principales: A, B y O. Un individuo solo puede poseer dos de ellos, pero en la población pueden coexistir varias combinaciones, lo que da lugar a una gama más amplia de fenotipos.
4.4. Epistasis e interacciones génicas
La epistasis se produce cuando la expresión de un gen depende de la presencia o ausencia de alelos en otro gen distinto, alterando o enmascarando su manifestación fenotípica. Este fenómeno evidencia la compleja red de interacciones génicas que existe en los organismos y que muchas veces impide predecir fácilmente el fenotipo a partir del genotipo. De ahí que la herencia real de muchos caracteres sea más compleja que la simple dicotomía dominante-recesivo.
4.5. Herencia ligada al sexo
Algunos rasgos se heredan de manera ligada a los cromosomas sexuales (X o Y). Esto implica que la dominancia o recesividad puede manifestarse de manera distinta en varones y mujeres, sobre todo en aquellos genes ubicados en el cromosoma X. Un ejemplo típico es el daltonismo ligado al cromosoma X. Como los varones tienen solo un cromosoma X, basta con que hereden el alelo mutado para mostrar la condición. En cambio, las mujeres requieren dos copias del alelo recesivo para manifestar el daltonismo, debido a que poseen dos cromosomas X.
5. Ejemplos de rasgos dominantes y recesivos en humanos
5.1. Color de ojos
El color de ojos en humanos es un rasgo poligénico (varios genes contribuyen al fenotipo), pero tradicionalmente se ha simplificado diciendo que los ojos oscuros (marrones) son dominantes sobre los ojos claros (azules o verdes). Esta simplificación no refleja toda la complejidad genética implicada, pero sirve para ilustrar el concepto de dominancia. En realidad, existen múltiples loci y alelos que interaccionan para dar lugar a la amplia gama de colores de ojos que encontramos en la población humana.
5.2. Grupo sanguíneo ABO
En este sistema, los alelos A y B suelen considerarse co-dominantes, mientras que el alelo O es recesivo. De esta forma, un individuo con genotipo AO manifestará el grupo sanguíneo A, mientras que quien tenga BO manifestará el grupo B. Por su parte, si un individuo hereda los alelos A y B, exhibirá el fenotipo AB. Finalmente, el individuo con OO presentará el grupo O, cuya proteína superficial no está presente en la membrana de los eritrocitos.
5.3. El hoyuelo en la barbilla
Algunas características faciales se han descrito siguiendo patrones mendelianos simples. El hoyuelo en la barbilla suele describirse como un rasgo dominante, aunque la expresión puede variar en diferentes poblaciones. No obstante, la genética moderna tiende a matizar estos ejemplos, destacando que hay muchos genes involucrados en el desarrollo facial, por lo que no es raro observar variabilidad en la expresión de estos rasgos.
5.4. Dimples (hoyuelos en las mejillas)
Los hoyuelos en las mejillas se han considerado tradicionalmente rasgos dominantes, pero existen numerosas excepciones que indican que otros factores genéticos y ambientales pueden modular su aparición. Aun así, siguen utilizándose como ejemplo clásico en la enseñanza de la genética a nivel escolar para ilustrar cómo un “rasgo dominante” puede expresarse en la descendencia, incluso si solo uno de los progenitores lo posee de forma visible.
5.5. Capacidad de enrollar la lengua
La habilidad para enrollar la lengua (tongue rolling) se ha mencionado a menudo como un rasgo dominante. Sin embargo, investigaciones más recientes sugieren que no se trata de un rasgo puramente mendeliano, pues hay factores adicionales que determinan esta capacidad. Este ejemplo es muy ilustrativo de la complejidad de la herencia en humanos y de cómo los ejemplos clásicos en ocasiones simplifican en exceso la realidad biológica.
6. Mecanismos moleculares de la dominancia y recesividad
6.1. Producción de proteínas y su relación con el fenotipo
Para entender por qué un alelo se considera dominante o recesivo, se debe analizar la cantidad y la funcionalidad de la proteína producida. En general, el alelo dominante produce una proteína funcional que determina el fenotipo. El alelo recesivo, en muchos casos, puede generar una proteína no funcional o no producirla en absoluto, de modo que si el organismo cuenta con una sola copia de un alelo dominante que cumpla la función requerida, se manifestará el fenotipo asociado a dicho alelo.
6.2. Haplosuficiencia
El término haplosuficiencia hace referencia a la situación en la que tener una sola copia funcional de un gen (en un individuo heterocigoto) es suficiente para producir el fenotipo normal o “dominante”. Cuando esto ocurre, hablamos de dominancia completa, porque el alelo recesivo no se expresa en el fenotipo. Por el contrario, si una sola copia funcional no es suficiente para alcanzar el fenotipo normal, el rasgo puede presentarse de manera intermedia (dominancia incompleta) o surgir otros efectos fenotípicos.
6.3. Mutaciones de ganancia de función
Algunos alelos dominantes se deben a mutaciones de ganancia de función, en las que el producto génico adquiere una actividad mayor o diferente de la normal. Ejemplos de esto se observan en ciertas enfermedades genéticas, donde el alelo mutante ocasiona un fenotipo dañino a pesar de la presencia de un alelo normal, precisamente porque dicha mutación produce una proteína hiperactiva o tóxica.
6.4. Mutaciones negativas dominantes
También existen mutaciones denominadas negativas dominantes, en las que la proteína alterada interfiere con la función de la proteína normal, secuestrándola o inactivándola. Este fenómeno puede ocurrir en proteínas que actúan como complejos multimerícos, de modo que la proteína mutante forma parte del complejo y altera su funcionalidad. En estos casos, el individuo heterocigoto expresa el fenotipo de la mutación dominante, aun teniendo un alelo normal.
7. Influencia del ambiente y penetrancia
7.1. Efecto del ambiente en la expresión génica
La expresión de los rasgos hereditarios no depende exclusivamente de los genes. El ambiente también juega un rol fundamental al interactuar con la dotación genética de cada organismo. Factores como la nutrición, la temperatura, la exposición a sustancias químicas y otros elementos pueden modificar la expresión de ciertos genes. En algunos casos, un individuo con un genotipo determinado puede no expresar el fenotipo esperado debido a condiciones ambientales específicas.
7.2. Penetrancia y expresividad
La penetrancia se refiere al porcentaje de individuos con un genotipo concreto que muestran el fenotipo esperado. Si la penetrancia es completa, todos los individuos con el genotipo en cuestión expresan el rasgo asociado. Si la penetrancia es incompleta, solo un porcentaje de ellos lo hacen. La expresividad, por su parte, describe el grado o la intensidad con que se manifiesta un rasgo en aquellos individuos que lo presentan. Ambos conceptos subrayan que la genética no opera en el vacío, sino que los genes interactúan con el medio interno y externo.
8. Aplicaciones prácticas en medicina y biotecnología
8.1. Diagnóstico genético
La distinción entre alelos dominantes y recesivos es crucial en el diagnóstico de enfermedades genéticas. Por ejemplo, patologías autosómicas dominantes como la enfermedad de Huntington requieren que el individuo herede solo un alelo mutado para manifestar la enfermedad, mientras que en trastornos autosómicos recesivos como la fibrosis quística se necesitan dos copias mutadas para que el fenotipo patológico aparezca. Los análisis genéticos, que pueden incluir pruebas de cribado o secuenciación, ayudan a determinar el riesgo de heredar y transmitir estas enfermedades.
8.2. Consejería genética
En la consejería genética, profesionales de la salud especializados orientan a las familias sobre la probabilidad de transmitir o presentar ciertos rasgos o enfermedades genéticas. Conocer si un rasgo es dominante o recesivo es esencial para estimar las probabilidades de herencia y para tomar decisiones informadas sobre la reproducción. Además, la comprensión de las distintas formas de dominancia y la existencia de rasgos ligados al sexo o poligénicos permite ofrecer pronósticos más precisos.
8.3. Terapia génica y edición genética
En la actualidad, con el advenimiento de herramientas de edición genética como CRISPR-Cas9, se ha abierto la posibilidad de corregir mutaciones en genes que causan enfermedades. En casos de mutaciones recesivas, bastaría con restaurar una copia funcional del gen para que el paciente recupere la expresión de la proteína normal, dado el principio de haplosuficiencia. En mutaciones dominantes de ganancia de función o negativas dominantes, el enfoque puede ser distinto, ya que la solución podría pasar por inactivar el alelo mutante o sustituirlo por uno normal.
8.4. Mejora de especies en agricultura y ganadería
El entendimiento de la dominancia y recesividad tiene implicaciones directas en los programas de mejoramiento genético de plantas y animales de granja. Se buscan cruces que promuevan la aparición de rasgos deseables (por ejemplo, mayor rendimiento en cultivos o mayor resistencia a enfermedades en ganado) y se analizan cuidadosamente los genotipos de las líneas parentales para maximizar la probabilidad de obtener la descendencia óptima. Aunque las técnicas de biología molecular y la selección genómica han modernizado este proceso, la genética mendeliana sigue siendo una referencia básica.
9. Aspectos éticos y sociales de la manipulación genética
9.1. Selección de embriones
La posibilidad de seleccionar embriones libres de mutaciones recesivas o dominantes ha suscitado debates éticos profundos. La disponibilidad de diagnósticos genéticos preimplantacionales plantea la pregunta de hasta dónde es lícito intervenir para prevenir enfermedades genéticas. Al mismo tiempo, se discute el riesgo de que estas técnicas se utilicen para seleccionar rasgos no estrictamente relacionados con la salud, como podría ser el color de ojos o características físicas.
9.2. Edición genética en humanos
La edición genética de línea germinal, que implica modificaciones que se transmitirán a la descendencia, se encuentra en un área altamente controvertida. Por un lado, ofrece la esperanza de erradicar enfermedades hereditarias al corregir mutaciones dominantes o recesivas. Por otro lado, se teme que el mal uso de estas tecnologías conlleve prácticas eugenésicas o desigualdades sociales, en un escenario en el que solo unos pocos tendrían acceso a estos avances.
9.3. Conservación de la biodiversidad
En el ámbito de la conservación, entender la genética de poblaciones y los rasgos dominantes y recesivos es fundamental para diseñar estrategias que mantengan la diversidad genética de especies amenazadas. Evitar cruces consanguíneos y preservar la variabilidad alélica contribuye a la resiliencia de las poblaciones frente a cambios ambientales y enfermedades.
10. Ejemplos experimentales y estudios de caso
10.1. Mus musculus (ratón de laboratorio)
El ratón de laboratorio es un modelo muy común en investigaciones genéticas. En estos roedores, se han identificado numerosos genes que muestran patrones de dominancia y recesividad. Experimentos para estudiar enfermedades humanas se basan en la introducción o supresión de genes específicos, evaluando el efecto en el fenotipo y comprobando si la mutación se comporta como dominante o recesiva. Este conocimiento ha sido vital para el desarrollo de medicamentos y terapias.
10.2. Drosophila melanogaster (mosca de la fruta)
Drosophila melanogaster es un organismo modelo clásico en genética. Gracias a su corto ciclo de vida y a la facilidad de manipulación en el laboratorio, se ha empleado para desentrañar mecanismos moleculares y patrones de herencia. Muchos de los genes estudiados en Drosophila tienen homólogos en mamíferos, incluidos los humanos. Además, la mosca de la fruta ha sido fundamental para descubrir la existencia de herencia ligada al sexo, epistasis y otros patrones que van más allá de la simple dominancia-recesividad.
10.3. Herencia de la pigmentación en plantas
En el mundo vegetal, el color de las flores y la pigmentación en las hojas son rasgos muy estudiados para comprender la interacción entre múltiples genes. Por ejemplo, en la flor de la boca de dragón (Antirrhinum majus), se ha investigado la presencia de múltiples genes que controlan la síntesis de antocianinas, pigmentos que dan color a los pétalos. En estos casos, se han descubierto fenómenos de dominancia incompleta, codominancia y epistasis, enriqueciendo el panorama de la herencia genética en plantas.
11. Tabla de ejemplos de rasgos dominantes y recesivos
A continuación, se presenta una tabla con ejemplos clásicos (y algunos discutidos) de rasgos en humanos que tradicionalmente se han descrito siguiendo el modelo mendeliano de dominancia y recesividad. Es importante recalcar que muchos de estos rasgos están sujetos a matices y no siempre se comportan de manera estrictamente mendeliana.
| Rasgo | Alelo Dominante | Alelo Recesivo | Comentarios |
|---|---|---|---|
| Color de ojos | Ojos oscuros (simplificación) | Ojos claros (simplificación) | En realidad, es poligénico. Hay múltiples genes involucrados. |
| Grupo sanguíneo ABO | A y B (co-dominantes) | O | Un individuo con alelos A y B presenta grupo AB. |
| Hoyuelos en mejillas | Presencia de hoyuelos | Ausencia de hoyuelos | Ejemplo clásico, pero con variaciones en la penetrancia. |
| Pico de viuda | Presencia de pico de viuda | Línea capilar recta | Se discute su pureza mendeliana; influido por varios factores. |
| Enrrollar la lengua | Capaz de enrollarla | Incapaz de enrollarla | Subjetivo y no puramente mendeliano, pero usado didácticamente. |
| Dimples en la barbilla | Con hoyuelo | Sin hoyuelo | Ilustrativo de la variabilidad de rasgos faciales. |
| Earlobe attachment (unión del lóbulo de la oreja) | Lóbulo libre | Lóbulo adherido | Ejemplo muy simplificado; hay considerable variación. |
12. Complejidades en la herencia humana
12.1. Poligenia
Muchos de los rasgos en humanos, como la estatura o la pigmentación de la piel, son poligénicos. Esto significa que dependen de la acción combinada de varios genes, cada uno con un pequeño efecto. Estos casos no se ajustan fácilmente al modelo mendeliano simple de dominancia y recesividad, ya que el resultado fenotípico es un gradiente continuo (como la variación en la estatura o en el tono de piel), en lugar de categorías discretas.
12.2. Herencia multifactorial
En la herencia multifactorial, tanto múltiples genes como factores ambientales contribuyen al fenotipo. Enfermedades comunes como la diabetes tipo 2, la hipertensión y la obesidad tienen una base genética compleja a la que se suma la influencia del estilo de vida, la dieta y otros condicionantes. En estos escenarios, no puede hablarse de dominancia o recesividad de un único gen, sino que hay redes génicas y contextos ambientales que determinan la aparición del rasgo o la patología.
12.3. Herencia mitocondrial
La mitocondria posee su propio ADN, diferente del ADN nuclear, y se transmite principalmente por vía materna en la mayoría de los seres eucariotas, incluidos los humanos. Cualquier mutación en el ADN mitocondrial afecta a los hijos de la madre portadora. Este tipo de herencia escapa completamente a la dicotomía de dominancia y recesividad, y se rige por mecanismos distintos relacionados con la fisiología mitocondrial.
13. Perspectivas futuras y líneas de investigación
13.1. Genómica personal
La secuenciación del genoma personal está cada vez más disponible a un coste reducido. Esto permite a las personas conocer sus variantes alélicas y su predisposición a diversas enfermedades, así como ciertos rasgos fenotípicos. El reto futuro es la interpretación funcional de estas variantes en un contexto poligénico y ambiental, lo que a su vez implicará un mejor entendimiento de los mecanismos de dominancia, recesividad y otras formas de interacción génica.
13.2. Epigenética y regulación génica
La epigenética estudia los cambios en la expresión génica que no se deben a alteraciones en la secuencia de ADN, sino a modificaciones en la cromatina, metilación del ADN, etc. Estos cambios pueden silenciar o activar genes de manera heredable. Incluso un alelo “dominante” puede silenciarse epigenéticamente en algunos contextos, lo que modifica los patrones de herencia más allá de la clásica perspectiva mendeliana. La epigenética es, por tanto, un campo de enorme relevancia para la comprensión integral de cómo se transmiten los rasgos.
13.3. Interacciones con el microbioma
Investigaciones recientes señalan que los microorganismos que habitan en nuestro cuerpo (microbioma) pueden influir en la expresión y el desarrollo de múltiples rasgos, incluidas algunas enfermedades. Si bien este ámbito de estudio está en expansión, no sería sorprendente descubrir que la presencia y proporción de ciertas bacterias modula la expresión de alelos recesivos o dominantes, añadiendo un nivel adicional de complejidad.
13.4. Ingeniería genética y organismos modificados
Las técnicas de ingeniería genética seguirán perfeccionándose y expandiendo sus aplicaciones. Además de la terapia génica en humanos, la modificación de organismos para fines agrícolas, industriales o médicos se está volviendo cada vez más sofisticada. El estudio de la dominancia y recesividad seguirá siendo esencial para predecir cómo se manifestarán los rasgos introducidos y cómo interactuarán con el fondo genético del organismo receptor.
Más Informaciones
